Ноцицептивные сигналы возникают в окончаниях первичных сенсорных нейронов и поступают в спинной мозг сначала по периферической, а затем центральной ветвям их аксонов. Теоретически биполярная структура первичного сенсорного нейрона допускает преобразование импульсных последовательностей на этом пути. Учитывая это, физиологи ввели понятие “сенсорная единица” (специфический рецептор в избирательной сенсорной системе и ноцицептор — в ноцицептивной), которое объединяет периферическую ветвь аксона первичного сенсорного нейрона (афферентное волокно), ее тканевое окончание (терминаль) и возможные претерминальные клетки-сателлиты, функционально связанные с терминалью. Сенсорную единицу можно представить как некий функциональный модуль, для которого входной сигнал — внешний стимул, а выходной — последовательность нервных импульсов в периферической ветви аксона первичного сенсорного нейрона.
Периферическая ноциология начала развиваться в XIX в., когда уровень техники был еще недостаточен для решения ее задач. В 1838 г. немецкий физиолог И.П.Мюллер предположил, что каждому ощущению, включая боль, соответствуют определенные специфические нервные волокна (в современных терминах — первичные сенсорные нейроны и ноцицепторы). В конце XIX в. гистолог М.Бликс обнаружил специализированные точки кожи для определенных видов стимулов, физиолог М.Фрей установил различную чувствительность малых участков кожи к точечным механическим стимулам [2, 3]. Эти работы привели к представлению о сенсорной гетерогенности кожи и послужили основой для формулирования теории специфичности боли. Согласно этой теории, существуют специализированные кожные нервные волокна, чувствительные окончания (ноцицепторы) которых возбуждаются только вредящими стимулами.
В XIX в. появилась и другая, альтернативная, теория боли — теория интенсивности, согласно которой защитный рефлекс и боль возникают при чрезмерном раздражении любого нерва. Сформулировал ее в 1895 г. немецкий врач А.Гольдшайдер, заметивший, что при некоторых заболеваниях происходит пространственное и временнOе суммирование ноцицептивных стимулов [4]. Теория интенсивности предполагает, что все сенсорные единицы выполняют двойную роль как специфических рецепторов (например, тепловых), так и ноцицепторов.
В XX в. обе конкурирующие теории были экспериментально проверены с помощью развитых к тому времени гистологических методов и новых электронных приборов. Английские гистологи школы Г.Уэдделла не выявили связи между структурой и функцией нервных окончаний кожи человека [5]. Оказалось, что все разнообразие кожных ощущений передается нервными окончаниями всего двух типов — инкапсулированными и свободными. Диссонанс между гаммой кожных ощущений и скудным набором структурных типов сенсорных терминалей особо заметен в волосистой коже, где инкапсулированных терминалей почти нет. Гистологи заметили и другой важный факт: большинство микроскопических участков кожи снабжено свободными нервными окончаниями различных аксонов. Это означает, что даже строго локализованный стимул одновременно возбуждает различные сенсорные единицы.
Результаты гистологических исследований послужили основанием как для критики теории специфичности, так и для развития теории интенсивности, что привело к созданию теории паттерна (нейроимпульсного узора) Дж.Нейфа [6]. Согласно этой теории, ноцицептивному (как и любому другому) стимулу соответствуют определенные импульсные последовательности (паттерны) в группах нервных волокон. Как и теория интенсивности, теория паттерна не предполагает существования специализированных ноцицепторов для распознавания повреждающих стимулов. Считается, что одно и то же волокно выступает в разных ипостасях, внося вклад в суммарный импульсный узор, приводящий к тому или иному ощущению.
К сожалению, структурную однородность нервных окончаний гистологи без каких-либо оснований распространили и на их функциональную однотипность, поэтому критика теории специфичности оказалась некорректной. В действительности же внешне одинаковые нервные окончания могут резко отличаться по реакции на различные воздействия. Однако выяснилось это, лишь когда удалось зарегистрировать сигналы в нервных волокнах.
|
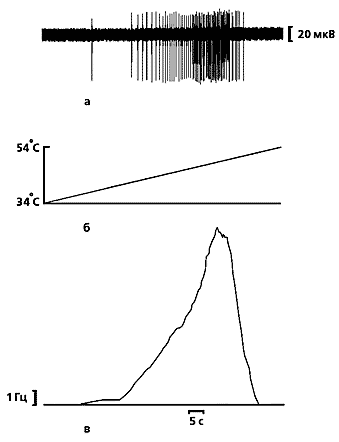
Типичная запись разрядов кожного ноцицептора (а) кошки при линейно-нарастающем нагреве (б), полученная в наших опытах [15]. Кривая (в) показывает среднюю частоту разрядов и отражает характерные особенности ноцицептивных сигналов индивидуальных С-ноцицепторов, которые представляют собой ограниченную последовательность из нескольких десятков нервных импульсов, следующих с частотой, не превышающей 20 Гц. |
|