


 Ранние исследования влияния pH на кислород-связывающие свойства крови миноги допускали, что фактор Бора мал (примерно от -0,1 до -0,3). Однако в тех исследованиях сродство гемоглобина к кислороду рассматривалось как функция от внеклеточного pH, поскольку в то время уникальность регуляции внутриклеточного pH в эритроцитах миноги не была известна. Более поздние исследования, в которых изменение сродства к кислороду соотносили с внутриклеточным pH показали, что фактор Бора достаточно велик. У Petromyzon marinus при 10° С и pHi=7.3-7.6 значение фактора Бора (DlogP50/DpHi) оказались равны –0,63, а в Lampetra fluviatilis при 18°С и при pHi=6,7-8,0 фактор Бора равен 1,03. Для Lampetra fluviatilis это значение показывает освобождение одного протона на один мономерный гемоглобин. Подобное же значение было получено на основе небикарбонатной буферной способности эритроцита, внутриэритроцитарной концентрации гемоглобина и внутриклеточных изменений pH при оксигенации. Таким образом эффект Бора гемоглобина миноги происходит из-за выпуска одного протона на одну цепочку гемоглобина при оксигенации.
Ранние исследования влияния pH на кислород-связывающие свойства крови миноги допускали, что фактор Бора мал (примерно от -0,1 до -0,3). Однако в тех исследованиях сродство гемоглобина к кислороду рассматривалось как функция от внеклеточного pH, поскольку в то время уникальность регуляции внутриклеточного pH в эритроцитах миноги не была известна. Более поздние исследования, в которых изменение сродства к кислороду соотносили с внутриклеточным pH показали, что фактор Бора достаточно велик. У Petromyzon marinus при 10° С и pHi=7.3-7.6 значение фактора Бора (DlogP50/DpHi) оказались равны –0,63, а в Lampetra fluviatilis при 18°С и при pHi=6,7-8,0 фактор Бора равен 1,03. Для Lampetra fluviatilis это значение показывает освобождение одного протона на один мономерный гемоглобин. Подобное же значение было получено на основе небикарбонатной буферной способности эритроцита, внутриэритроцитарной концентрации гемоглобина и внутриклеточных изменений pH при оксигенации. Таким образом эффект Бора гемоглобина миноги происходит из-за выпуска одного протона на одну цепочку гемоглобина при оксигенации.
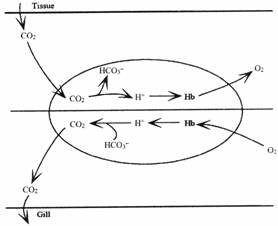
У миноги эффекты Бора и Хэлдена ориентированы на транспорт CO2. Эффект Хэлдена в интактных эритроцитах миноги настолько велик, что внутриэритроцитарный pH венозной крови выше, чем артериальной. Роль эффекта Хэлдена в транспорте CO2 кровью миноги становится ясной, если рассмотреть механизм этого транспорта. Вследствие относительной непроницаемости мембраны эритроцитов миноги для бикарбоната и других кислотно-основных эквивалентов, карбоангидраза, содержащаяся в эритроците недоступна для бикарбоната плазмы. Вследствие этого только небольшой процент бикарбоната плазмы может быть превращен в CO2 в некатализируемой реакции дегидратации, когда кровь проходит через жабры, потому, что полупериод некатализируемой реакции равен 1 минуте при 15 °С, а время пребывания крови в жабрах всего несколько секунд. Таким образом, большинство удаляемого из крови CO2 в жабрах приходит из эритроцитарных бикарбонатных запасов и эффективность транспорта CO2 зависит от того, какое его количество эритроциты смогут загрузить в тканях и выгрузить в жабрах. Главные факторы, влияющие на это: изменение давления CO2 в кровообращении и кривые диссоциации CO2. На основании кривых диссоциации CO2 для деоксигенированных и оксигенированных эритроцитов Lampetra fluviatilis понятно, что более 80 % уменьшения содержания CO2 в эритроцитах может быть приписано к зависящему от оксигенации уменьшению содержания CO2 и менее чем 20 %-к уменьшению давления CO2. Оксигенация не влияет на содержание CO2 в плазме. Зависящее от оксигенации уменьшение внутриэритроцитарного содержания CO2 полностью объясняется зависящим от оксигенации уменьшением внуриэритроцитарного pH, вызванным эффектом Хэлдана. Таким образом, основываясь на данных in vitro можно сказать, что эффекты Бора и Хэлдана определяюще влияют на транспорт CO2 кровью миноги. Хотя данные in vivo немного отличны, они так же показывают ведущую роль эритроцитов миноги в экскреции CO2 в сравнении: только 15 % выводимого CO2 у радужной форели происходит из эритроцитов.
Из-за высокого венозного внутриэритроцитарного pH (на 0.15-0.2 ед. выше, чем в артериальной) у Petromuzon marinus CO2 эффективно загружается в эритроцитарные запасы, в основном в виде бикарбоната. Вследствие уменьшения давления CO2 крови в жабрах, а так же, вследствие уменьшения внутриэритроцитарного pH из-за высвобождения протонов благодаря оксигенации гемоглобина, CO2 интенсивно выгружается в жабрах. Таким образом, уменьшение общего содержания CO2 при прохождении крови через жабры у миноги подобно таковому у активных костных рыб, таких как радужная форель, за исключением того факта, что бикарбонат плазмы у миноги играет очень ограниченную роль в выведении CO2 из-за медленного проникновения бикарбоната через мембрану.