


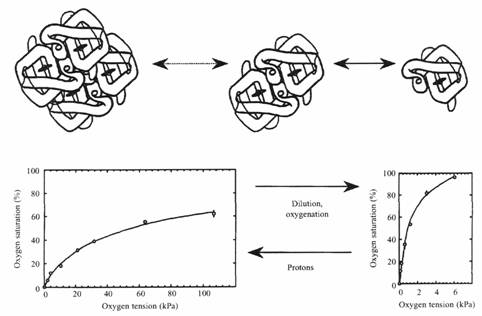
В разбавленных растворах сродство к кислороду гемоглобина миноги заметно зависит от его концентрации. Увеличение концентрации уменьшает сродство благодаря возрастанию процента низкосродственных олигомеров. Так как гемоглобин в интактных эритроцитах показывает кооперативность и эффект Бора, вероятно, реакции ассоциации\диссоциации гемоглобина протекают также и в эритроците. Если это имеет место, то сродство гемоглобина к кислороду должно зависеть от изменения клеточного объема (которое влияет на внутриклеточную концентрацию гемоглобина). Эта зависимость была проверена экспериментально путем осмотического сжатия клеток и последующего измерения P50. Результаты показали уменьшение сродства с уменьшением объема клетки. Кроме этого уменьшения сжатые эритроциты обычно имеют более высокий pH, чем эритроциты с нормальным объемом. Таким образом, концентрационные изменения оказываются способными вызывать изменения в агрегации молекул гемоглобина и, следовательно, сродства гемоглобина к кислороду. Пока не ясно, являются ли подобные изменения объема частью нормальной физиологической регуляцией функции гемоглобина, однако данные об адаптировавшихся к гипоксии Lampetra fluviatilis показывают, что средняя концентрация гемоглобина в эритроците становится ниже, после недели адаптации к гипоксии, чем при нормоксии.
Ограничение буферных свойств гемоглобина вследствие низкой проницаемости мембраны эритроцитов миноги к кислотным эквивалентам.
 |